DIRECT AND INDIRECT METHODS OF GENE TRANSFER IN PLANT
Plants are important sources of many important products, such as food, fibers, medicines, energy which benefits the need of human beings. Human has been cultivating these plants to meet their desired products. Theses selective plants were improving for better quality and quantity of product by breeding to meet the need of growing human population on Earth. The plant breeders are depended on the existing gene pool and sexual compatibility of the plant species, which is a limitation.
The genetic material of species were altered by the incorporation of the foreign DNA following the molecular biology methods are called transformation and if these techniques performed directly or indirectly in the plant cells are called plant genetic transformation. In the year 1981, the first successful gene transformation was demonstrated in the tobacco plant using the soil bacterium Agrobacterium tumefaciens. Until now more than 100 different plant species has been transformed with the desired foreign genes using A. tumefaciens or other available methods.
The plant genetic transformation has become a versatile method for the production of agricultural and medicinal value product for benefits of human society. The transfers of gene in to plants cells are difficult because plant cells are impermeable which acts as barrier to diffuse through cell membrane. Due to diversity of plant species and their diverse genotypes variety of gene transformation methods has been developed to overcome the barrier in plants.
PLANT GENETIC TRANSFORMATION METHODS
The various available plant transformation methods has been discussed in this chapter such as biological methods; Agrobacterium tumefaciens and protoplast mediated transformation, Chemical methods; calcium phosphate, co-precipitation and lipofection, physical methods, electroporation, biolistics, agitation with glass beads, vacuum infiltration, silicon carbide whisker, laser microbeams, ultrasound, and shock-wave mediated.
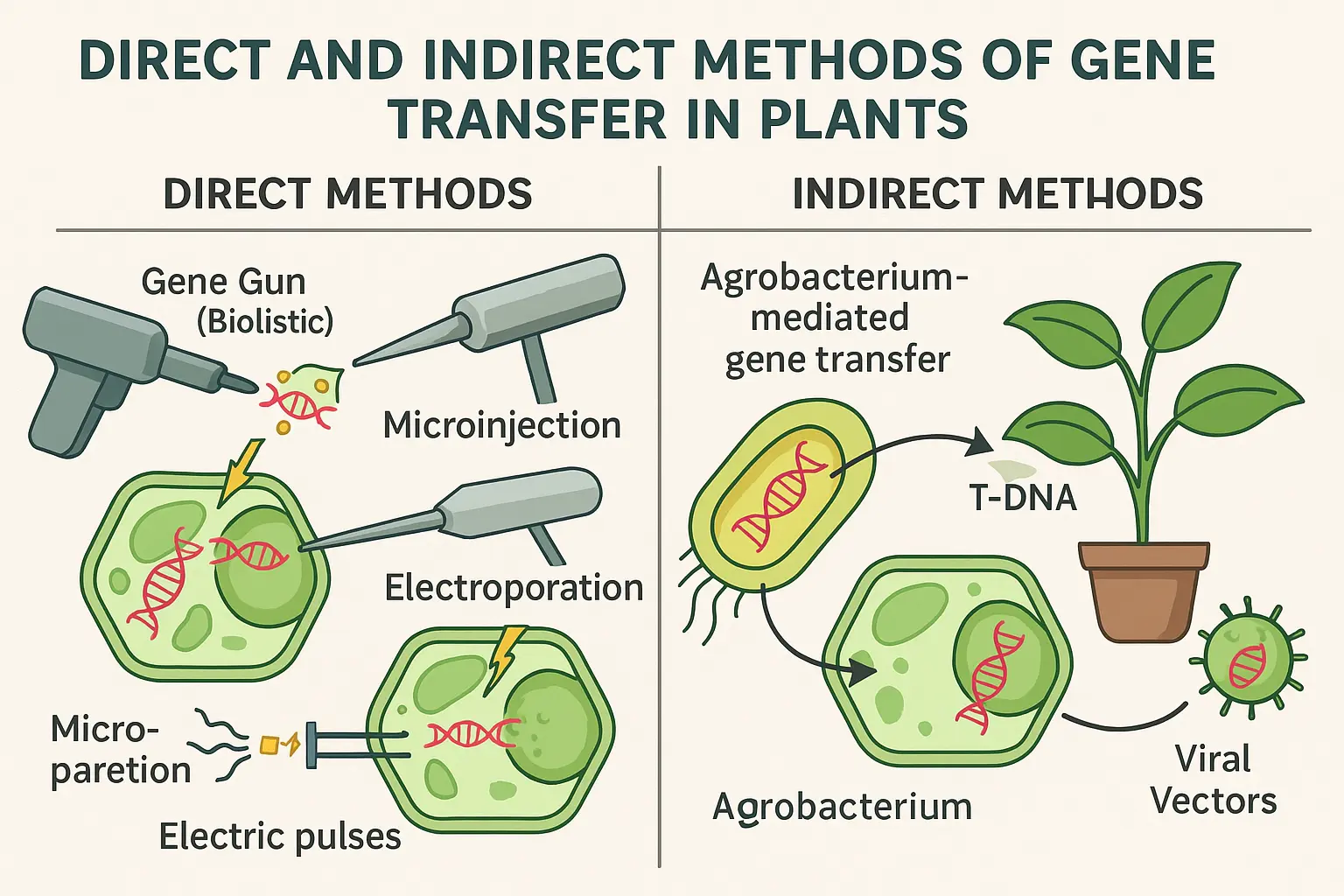
The plant genetic transformations are classified into direct and indirect gene transfer methods.
Indirect gene transfer methods
For indirect gene transformation methods, two bacterial strain, Agrobacterium tumefaciens, or Agrobacterium rhizogenes, has been discover to transfer the desired gene into plant cells however Agrobacterium tumefaciens is widely used.
Agrobacterium tumefaciens
Among the various available vectors for plant transformation the Ti plasmid of Agrobacterium tumefaciens has been widely accepted. Agrobacterium was first discovered in grape plant in the year 1897 by Fridiano Cavara. In 1907, crown gall disease was reported in plant caused by T-DNA of Ti-plasmid of Agrobacterium. Agrobacterium has been characterized as gram-negative bacteria or soil phytopathogen that belongs to the rhizobiaceae family. Agrobacterium prefer to infect mostly the dicotyledonous plants and at infection or wound site produces an unorganized growth of cells that known as crown gall tumors. This bacterium harbors the tumor inducing plasmid known as Ti plasmids which are exploited to transfer desired gene into target plant tissue. Agrobacterium got the natural ability to transfer the T-DNA, part of Ti plasmid, into the plants genome and due to this unique ability, Agrobacterium known as natural genetic engineer of plants. The unwanted sequence of T-DNA region of Ti-plasmid is the crucial region to replace with foreign desire gene.
Molecular basis of Agrobacterium-mediated transformation:
Vectors for gene transfer
Vectors usually contain the selectable markers to recognize the transformed cells from the untransformed cells, multiple rare restriction sites and bacterial origins of replication (e.g. ColE1). However, these features in vectors do not help in transfer of gene and integration into plant nuclear genome. Ti-plasmid of Agrobacterium has wide host range and capable to transfer gene that make them preferred vector over other available vectors.
The T-DNA nucleotide sequence end are flanked by 25 bp direct repeat sequences known as left border (LB) and right border (RB) and both border sequences collectively known as T-DNA border. Plasmid DNA comprising the T-DNA with border sequences called as mini or micro Ti-plasmid. Nucleotide sequences of T-DNA borders are essential and play an important role to transfer the T-DNA into the plant cell upon infection if present in cis orientation but border sequence itself does not get transfer. Any DNA sequence flanked by repeat of 25 bp in the correct orientation can be transfer to plant cells and similar attribute were exploited with Agrobacterium mediated gene transfer to produce transgenics of higher plants. It was shown in experiment only right border sequence have been used and observed an enhance sequence or sometimes refer overdrive sequence located upstream to right border sequence also necessary for high-efficiency transfer of T-DNA. However, the left-border sequence has little activity alone.
b. Structure and functions of Ti Plasmids
Agrobacterium harbor a large Ti-plasmid of 200-800 kbp which contains four main regions; T-DNA, vir region, origin of replication, region enabling conjugative transfer and o- cat region. The molecular understanding of crown gall disease caused created an opportunity to develop the gene transfer system in plant.
T-DNA:
It has been demonstrated that T-DNA of Ti-plasmid of Agrobacterium transfer to plant nuclear genome that cause the crown gall disease. If the Agrobacterium killed with antibiotics then also undifferentiated callus can be cultivated in in vitro retaining the tumorous properties. This property represents the oncogenic transformation of crown gall tissues and has the ability to form tumor if grafted onto a healthy plant. T-DNA, a small and specific element of Ti plasmid are of ~ 24kbp size which comprises the following important regions, (i) two tms genes responsible for indole acetic acid (an auxin) biosynthesis and tmr gene responsible for isopentyladenosine 5′-monophosphate (a cytokinin) synthesis. This is the reason when T-DNA sequence transferred to the plant nuclear genome leads to form crown gall due to the synthesis of two phytohormones, auxin and cytokinin (ii) os region responsible for synthesis of unusual amino acid or sugar derivatives, known as opines. Opines metabolism is the chief feature of crown gall disease formation. Two common opines, octopine and nopaline, synthesized in the plant cells from octopine and nopaline synthase, respectively. Agrobacterium strain determine the type of opine to be produce and not by host plant. Based on type of opine produced, Ti-plasmid further described as octopine type Ti – plasmid or nopaline type Ti – plasmid
Virulence gene
Vir gene is also essential for T-DNA transfer and unlike border sequence of T-DNA vir gene can function even in trans orientation. T-DNA and vir gene present on two different plasmids does not affect the T-DNA transfer provided both are present in same Agrobacterium cells. The virulence region of Ti-plasmid are of approximately 35 kbp in size and organized in eight operons known as Vir A, Vir B, Vir C, Vir D, Vir E, Vir F, Vir G and Vir H. All operon except Vir A and Vir H are polycistronic in nature.
Origin of replication:
In general, the Agrobacterium has broad host range of origin of replication for multiplication in different type of host plant.
Mechanism of T-DNA transfer to the plant genome:
The foreign gene transfer using the Agrobacterium has been considered a highly improved form of bacterial conjugation. The complete Ti-plasmid does not transfer to plant instead only a small segment or T-DNA get transferred and integrated into the plant genome. Virulence (vir) genes responsible for T-DNA transfer are located in a separate part of Ti- plasmid. T-DNA carries the genes for unregulated growth and also to synthesize opines in the transformed plant tissues. These genes are not necessary to transfer the T-DNA and therefore this region generates an opportunity to be replaced with the desired foreign genes.
Recognition and induction of vir genes: Plants secrets number of chemotactic signal or phenolic compounds but acetosyringone and β-hydroxyacetosyringone particularly and certain monosaccharides are recognized to induce the vir genes expression, processing, transfer and may also for integration of T-DNA into the plant genome. The bacteria respond well to the simple molecules like sugars and amino acids but not to the acetosyringone compounds to the injured plant cells. The vir genes induce only after attachment of agrobacterium to the plant cells. The chemotactic signal turns on Vir A by autophosphorylation and it will phosphorylates Vir G genes. Vir A and G gene expressed constitutively at low level. Vir A and G product encodes a membrane-bound sensor kinase and cytoplasmic regulator protein, respectively. It is well established that the signal transduction process involves Vir A autophosphorylation and then subsequently transfer phosphate to Vir G. Mainly vir G gene along with additional gene on the Agrobacterium chromosome encode transcriptional activator they play an important role to regulate the other vir genes. The list of vir genes of Ti-plasmid of Agrobacterium and their function are summarized in Table 1.Table 1. List of Vir locus of Ti-plasmid of Agrobacterium and their function.
Transfer of T-DNA to plant cells: Transfer of T-DNA initiated by Vir D1 and D2 product an endonuclease enzyme which specifically recognizes T-DNA border sequences. These enzymes create nick either on single strand or double strand at T-DNA border sequences which leads to release of ss-T-DNA from the Ti-plasmid. This process is enhanced by the Vir C2 and 12 proteins by recognizing and binding to the enhancer elements. Vir D2 binds covalently to 5’end of processed ss T-DNA forming an immature T-DNA complex. Single stranded T-DNA intermediate favored by the octopine type Ti-plasmid whereas double stranded favored by nopaline type Ti–plasmid. Vir D2 protein protects the T-DNA intermediate complex from nuclease degradation to target the DNA to cytoplasm and nucleus then integrate into plant genome. T-DNA intermediate are coated with Vir E2 which encodes a single stranded DNA binding protein (SSBP). The induction of vir gene expression forms the conjugative pilus to transfer the T-DNA to plant cells. Vir B gene operon product also involved to make part of conjugative pilus. Vir B and Vir D product then transport the T- DNA complex through membrane channel a type IV secretion system to cytoplasm of plant cell. Vir D4 protein acts as a linker facilitating the interaction of processed T-DNA complex with membrane channel. Vir B2 to Vir B11 and Vir D product are important for forming a membrane-associated export apparatus includes hydrophobicity, membrane-spanning domains, and/or N-terminal signal sequences in cytoplasm. The interaction between Vir B7 and Vir B9 help to form heterodimer that stabilizes the other Vir B proteins.
Membrane channel is composed of Vir D4 and Vir B11 protein which is necessary for transport of T- DNA complex. Vir B protein also serves as ATPases to provide energy for channel assembly or export process. Vir B1 has transglycosidase activity which utilizes to assemble other Vir proteins. With the T-DNA complex the other Vir protein H and F also transported into plant cells which are necessary for efficient transport of T-DNA complex and nuclear transport. Next, Vir D2 and Vir E2 protein play an important role for nuclear transport of T-DNA complex because it contains the nuclear localization signal. The nucleus of injured plant cells often associated with the cytosolic membrane facilitating the rapid transfer of T-DNA into nucleus without much exposure to the cytosolic environment. After reaching to the nucleus, T-DNA probably integrate to plant genome by illegitimate recombination process exploiting naturally occurring chromosome breaks.
Ti-plasmid derivatives for gene transfer
T-DNA of Ti-plasmid transferred and express in to plant cells because T-DNA carry the promoter element and polyadenylation site similar to eukaryotic one. This sequence acquired by agrobacterium may be during the evolution of Ti-plasmid. Ti-plasmid of Agrobacterium can transfer its T-DNA to the plant genome due to this specific reason Ti- plasmid qualified as natural vector to engineer the plant cells. The wild type Ti-plasmid is suitable due to presence of oncogenes on the T-DNA which causes the uncontrolled growth of plant cells. So the oncogenes region of T-DNA must disarmed to be qualified as successful natural vector for revival of plant well. T-DNA should be left with the left- and right border sequence and the nos gene in modified vector. The plant cells when transform with Agrobacterium containing modified vector, no tumor should produce and nopaline production will be evident for positive transformation. To make the screening easier to identify the transformed plants cells, selectable marker like drug or herbicide resistance can be inserted on T-DNA because the enzymatic assay for nopaline at every step of transformation is a cumbersome process. Nopaline positive cells could be culture to callus tissues provided the required phytohormones.
The modified vector of Ti-plasmid is also not convenient for plant transformation due to the large size that makes them difficult for manipulation. The absence of unique restriction enzymes sites in T-DNA sequence is another problem to manipulate this vector.
This problem was resolved by constructing an intermediate vector, in which T-DNA was subcloned into E. coli plasmid vector for easy manipulation. However, intermediate vector does not replicate in Agrobacterium and also lacked conjugation functions. To succeed the gene transfer process the triparental mating was introduced mixing the three bacterial strains like: a) E. coli strain carrying the recombinant intermediate vector; b) E. coli strain containing the helper plasmid to mobilize intermediate vector in trans; c) Agrobacterium carrying the Ti- plasmid. Conjugation between E. coli strains- a and b, transferred them to the c, recipient Agrobacterium. Homologous recombination occurs between the T-DNA and intermediate vector and forms a large cointegrate plasmid, from where recombinant T-DNA transferred to plant genome. Intermediate vector has been widely used but large cointegrate vector still not required for gene transformation in plant.
T-DNA sequence is not essential and the necessary vir gene region can functions in trans during the transfer the gene by Ti-plasmid to the plant. Therefore, vir gene and disarmed T-DNA sequence part of Ti-plasmids can supply on separate plasmids in Agrobacterium and this principle were termed T-DNA binary vector system. In binary vector system, maintaining the T-DNA on a shuttle vector is beneficial because the copy number is not determined by Ti-plasmid and not dependent on recombination. This event makes the identification of transformats much easier. The gene of interest to transfer including origin of replication and antibiotic resistance genes will be maintained on T-DNA region in binary vector system, whereas vir gene maintained on separate replicon known as Vir helper plasmid. The vir gene products will help processing the T-DNA and export further to plant cells (Figure 1 & 2). Ti-plasmid with vir gene region but without the T-DNA sequence will be transformed into the Agrobacterium. T-DNA sequence will introduced in to Agrobacterium by triparental mating or methods like electroporation. To achieve the efficient transformation the binary vector should have some properties: a) right and left border sequence of T-DNA; b) Selectable marker gene compatible to plant usually antibiotic or herbicide resistance; c) Multiple rare-cutting restriction endonuclease site on T-DNA and the lacZ gene for blue-white screening and cos site for preparing cosmid libraries; d) Origin(s) of replication for E. coli and Agrobacterium facilitate the replication in broad host range; e) Antibiotic-resistance genes in binary vector for selection in both E. coli and Agrobacterium.
Agrobacterium-mediated gene transformation in plants
For the transfer of gene in dicot plants, a few millimeters diameter leaves were surface sterilized and inoculated in medium containing Agrobacterium cells transformed with recombinant disarmed binary or cointegrate vector. The leaf disk was first grown for two days and then transferred to the medium containing the kanamycin and carbenicillin. In the medium, kanamycin added because the foreign chimeric gene has kanamycin resistance gene for selection and carbenicillin to kill Agrobacterium cells. The shoots were usually developed in 2-4 week time from the leaf disk. The grown shoots were removed from callus and transfer to the medium containing auxin for root development. The roots were developed in 2-3 week time and then plantlets were transplanted to the soil. This is a superior, simple and rapid methods compared to methods where transformed plant were recovered from the protoplast-derived callus which transformed with agrobacterium by co-cultivation.
In case of monocot plants, only few of the monocots plant such as rice, maize, wheat, barley, and sugarcane were reported susceptible to Agrobacterium infection with the modified culture condition and transformation procedures. The use of explant, embryo and apical meristem and supervirulent strain of Agrobacterium like AGL-1 was the key factor for successful transformation. Transformation efficiency in rice was achieved by adding acetosyringone in co-cultivating medium of Agrobacterium and rice embryos.
Agrobacterium rhizogenes:
A. rhizogenes is another bacterial strain use to transfer the gene of interest to the plant cells. The molecular understanding of hairy root diseases helps to utilize this Agrobacterium for gene transfer system, analogous to A. tumefaciens. This Agrobacterium harbor the Ri- plasmid which are responsible to produce the characteristic hairy root disease symptoms upon infection to dicotyledonous plant. Ri-plasmid also has T-DNA region which transfer into the plant nuclear genome. T-DNA integrates into the plant genome and in turn iaaM and iaaH gene induced to produces excess phytohormones tryptophan 2-monooxygenase and indoleacetamide hydrolase, respectively. There are no major differences observed between the Ri plasmid and Ti-plasmid. However, it is not accepted commercially because of problem involved in scale-up of transformed roots.
Direct DNA transfer to plants
Direct gene transfer methods can also called as vector free methods because no living vector used to transfer gene to plant cells. Many physical and mechanical methods were developed that facilitate the entry of this foreign DNA into plant cells.
Protoplast Transformation
The protoplast cells are capable to take up the gene of interest from their surrounding liquid environment. After entry of gene of interest into protoplast, it gets integrated into the genome of transfected cells. The selectable marker can be also added with the gene of interest that required for the selection of desired gene in the protoplast. The gene transfer process can be induce and accelerated under influence of some chemicals like polyethylene glycol (PEG). Alternatively the electroporation methods can also be used for gene transfer to the protoplast. The putative protoplasts containing gene of interest were grown on selective medium where protoplast regenerate their cell wall, cell division begin and eventually produces the callus. Callus then produces the roots and shoots by inducing with phytohormones. The major problems often observe the regeneration of host plant from the protoplast however dicots found more responsive than the monocots.
Electroporation
Electroporation method was first demonstrated studying gene transfer in mouse cells and can be applied also with bacterial, fungal and plant cells. It is a simple and efficient method for integration of gene of interest into protoplast or intact plant cells. For electroporation, with high voltage (1.5 kV) short duration and with low voltage (350V) long duration of pulse was used for gene transfer. Electroporation pulse increases the permeability of membrane by disrupting the phospholipid bilayer of protoplast. This is in turn facilitating the entry of gene of interest into cells if present on protoplast membrane. The target cells can be pre-treated with enzymes or wounded for ease of gene transfer process. Without any form of pre-treatment also gene transfer has been successfully achieved in immature rice, maize and wheat embryos by electroporation method.
The efficient and successful gene transfer by electroporation methods depends on the following factor like applied electric field strength, electric pulse length, temperature, DNA conformation, DNA concentration, and ionic composition of transfection medium, etc. PEG can also be used to stimulate and enhance the uptake of liposome and also improve the transformation efficiency. Successful transfer of gene of interest using electroporation methods has been already achieved in maize, petunia, rice, sorghum, tobacco crops. Some new measure has been suggested to increase the transformation efficiency such as i) uses of 1.25kV/cm, ii) add first DNA followed PEG, iii) heat shock at 45°C for 5 minutes and iv) use linear DNA in place of circular. The modified condition are suitable to transfer gene of interest in both monocot and dicot protoplast.
Particle bombardment
Particle bombardment methods of gene transfer also known as biolistics, or particle gun, or gene gun, or short gun, or microparticle gun, or projectile bombardment (Figure 3) method. This method is especially useful when some of the live plant tissue like intercellular organelles, leaves, meristem, immature embryos, callus or suspension cultured cells, live pollen, which are impermeable to foreign DNA. The type of plant material used for DNA delivery is not a limitation in this method because the DNA delivery is governed by physical parameters. Plant cell wall is hard and not easy to delivers anything from outside so the powerful particle bombardment method is very useful for efficient gene transfer in plant. Particle bombardment method was developed at Cornell University. The gene of interest, DNA or RNA coated to tiny biologically inert high density particles like gold or tungsten of 1-3 pico to micrometer size are place on the target tissue in vacuum condition. Then gene coated high density particles are accelerated for high velocity (1400 ft/sec) by powerful shot using gene gun to enter inside the tissue membrane. The explosive charge like cordite explosion or shock waves initiated by high voltage can be used to get the high velocity acceleration in the gene gun. The success of particle bombardment method are governed by some factors such as particle size, acceleration (for penetration and determine the tissue damage), amount and conformation of DNA. These four factors must optimize for each species and type of target tissue using for gene transfer for success of gene transfer. Using this method the first successful transgenic were produced in soybean transferring gene to meristem tissues which were isolated from immature seeds. Gene gun method has been successfully used to transfer gene of interest in crops like barley, cotton, maize, oat, papaya, rice, soybean, sugarcane, tobacco, wheat. This method has also been used to transfer gene of interest for transient expression in onion, maize, rice and wheat.
Over the years the particle bombardment methods has been modified for better control over particle delivery, efficient transformation of gene and enhanced reproducibility of transformation states. For example, particle bombardment based on electric discharge has been designed for gene transfer in recalcitrant cereals and legume crops. Other gene gun modification includes like pneumatic apparatus, particle inflow gun using flowing helium and device utilizing compressed helium.
Microinjection
Microinjection is widely used and efficient technique for transfer of desire gene into animal cells, tissues or embryo cells nuclear genome. This technique is not efficient to direct transfer gene to the plant cells. In this method gene transfer to the cytoplasm or nucleus of recipient protoplast or plant cells were performed with the glass micropipette of 0.5-10μM diameter needle tip. The target recipient cells for gene transfer are many such as immature embryos, meristems, immature pollen, germinating pollen, isolated ovules, embryogenic suspension cultured cells, etc. The recipient cells were immobilized on a solid support like depression slide under suction and then the cell membrane and nuclear envelope of plant cells were penetrated with the glass micropipette tip under specialized micromanipulator microscope set up. The modified and improved method of microinjection termed as holding pipette method, in which the protoplasts positioned on a depression slide close to already placed micro-drop desired DNA solution. Now with the holding pipette hold the protoplast and DNA were injected into protoplast nucleus by injection pipette. Many genetic manipulation experiments were widely performed using this technique for cell modification, silencing of gene etc. This microinjection technique where also performed and demonstrated successful gene transformation and transient expression in green algae, Acetabularia. The drawback of this microinjection technique is that process is very slow, expensive, required highly trained technician and only a part of plant transformed with the desired gene. However, the success rate of transfer of gene is very high. This technique has been employed successfully in oilseed rape (Brassica napus) and obtained the transgenic chimera.
Sonoporation:
In this methods the explant like leaves are chopped into pieces and using the ultrasound waves creates the permeability function in cell wall. Through this permeable cell wall, the gene of interest can be uptake from surround environment by cell wall. Sonoporation process uses the sound waves which help to form tiny bubbles that enhance the DNA entry into cell walls. The explants were further transferred to the culture nutrient medium for the growth of shoots and roots.
Calcium phosphate mediated:
Calcium phosphate mediated gene transformation method was also considered promising for plant cells. In this method the desired gene with Ca2+ ions precipitated and forms calcium phosphate which coats the cells and released inside the plant cells. Using this method the desired gene can be transferred to study the molecular, biochemical, cellular, genomic and proteomic aspects in in vitro and in vivo of plant cells.
Lipofection:
Lipofection is a liposome-mediated gene transfer method. This method employs a liposome containing desired gene which induced by PEG to transfer of gene and then fuse into protoplasts. Liposome is cationic in nature and made up of phospholipid layer similar as cell membrane. Liposome and target cells adhere and form aggregates easily because of similar phospholipid bilayer (Felgner et al., 1987). The aggregate of liposome and cell wall are positively charge which enhances the efficiency of negatively charged DNA uptake. The desired gene enter into protoplast by endocytosis process of liposome that includes adhesion of the liposomes to the protoplast surface, liposomes fusion at the site of adhesion and then finally release of DNA inside the protoplast cell. There are many advantages with this method over other gene transfer method like desired gene not exposed to nuclease, stability due to encapsulation, low cell toxicity, high degree of reproducibility and suitable for wide range of cell types. Lipofection method of gene transfer has been successfully used in number of plant species like tobacco, petunia, carrot.
Join Times of Agriculture
Get the latest Agriculture Magazine and regular important updates right on your phone.
👉 Join WhatsApp Group